Place Cells: The Neurons That Know Where You Are
In 1971, John O'Keefe recorded from individual hippocampal neurons in freely moving rats and found cells that fired only at specific locations — place cells. This article explains what place cells are, how they form a cognitive map, what the 2005 discovery of entorhinal grid cells revealed about their upstream coordinate system, and why the place cell framework now extends far beyond spatial navigation into memory, episodic recall, and abstract cognition.

The experiment that changed how we understand memory and navigation
In 1971, John O'Keefe was trying to understand what the hippocampus actually did. The region had been linked to memory since the famous case of H.M. in the 1950s, and earlier rodent work had shown that hippocampal damage made rats terrible at spatial tasks. But "involved in memory and space" is a description, not a mechanism. O'Keefe wanted to see the neurons in action.
Together with his student Jonathan Dostrovsky, he implanted microelectrodes in the hippocampi of freely moving rats and recorded from individual neurons as each animal explored a small platform — just 24 cm by 36 cm. The setup was unusual. Most neuroscientists at the time recorded from animals held in fixed positions; following a rat around a maze while it did whatever it wanted was close to heresy. 1
Out of 76 hippocampal recording sites, eight behaved in a way that had no precedent. They fired — rapidly, reliably — only when the rat occupied a specific location on the platform, facing a specific direction. Move the rat somewhere else, and the cell fell silent. Return it to the same spot, and the cell fired again.
O'Keefe and Dostrovsky called their short 1971 report "The hippocampus as a spatial map." It was preliminary evidence — eight cells, a tiny platform — but the phrase contained a whole theory of how the brain represents the world. 2
What a place cell actually is
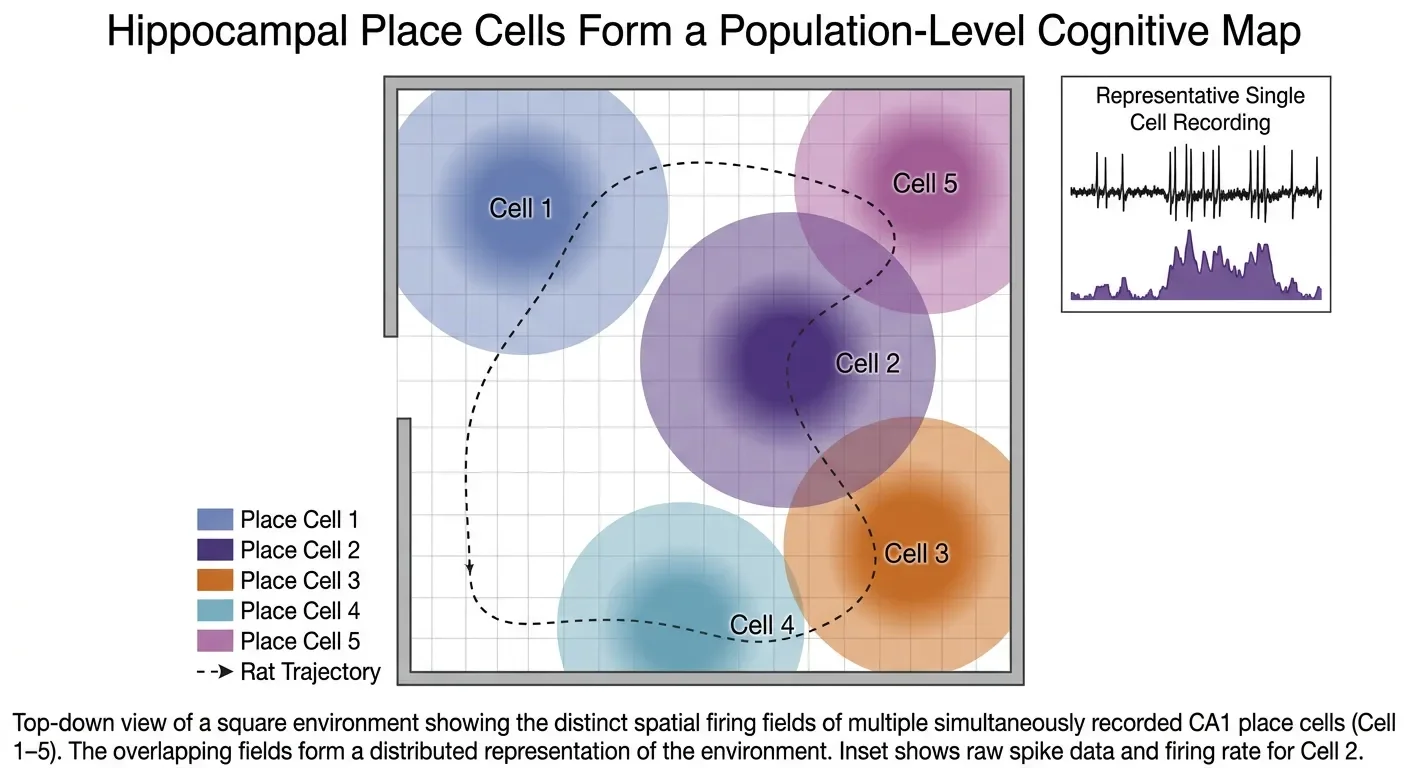
A place cell is a pyramidal neuron in the hippocampus — concentrated in the CA1 and CA3 subfields — that has a place field: a specific region of an environment where it fires. Outside that field, it sits quiet, producing fewer than one spike per second. Inside the field, it bursts at rates that can exceed 100 spikes per second. The field is allocentric, meaning it is anchored to the external environment, not to the animal's own body position. Two neighboring place cells can have place fields at completely opposite ends of a maze; there is no topographic order the way there is in, say, visual cortex or somatosensory cortex.
The precision is remarkable. A typical place field in a small environment covers roughly a tenth to a quarter of the available space. In larger environments, place cells may have multiple fields, and in truly large open areas the fields expand — consistent with the idea that the brain is fitting its representation to the scale of the available territory. 3
Place cells work as an ensemble. No single cell gives the animal its location; rather, the pattern of which cells are firing — the population vector — encodes position. At any given location, a small fraction of place cells are active and a large majority are silent. As the animal moves, the active population shifts, with cells at the animal's destination beginning to fire before the cells at the departure point have finished. The result is a continuous, overlapping representation of position as the animal traverses space.
The cognitive map theory
O'Keefe went further than just describing the cells. In 1976 he published a longer study, and in 1978 he and colleague Lynn Nadel published The Hippocampus as a Cognitive Map — a 600-page book that argued the hippocampus constructs and maintains an allocentric spatial representation of the environment, a cognitive map in the sense that the psychologist Edward Tolman had proposed in the 1940s. 4
Tolman had shown that rats could learn the layout of a maze even without direct reward, and could take novel shortcuts they had never been trained on — behavior that implied an internal spatial model, not just a learned chain of stimulus-response associations. O'Keefe and Nadel proposed that place cells were the physical substrate of that model.
The theory made a prediction that could be tested directly: if you destroy the hippocampus, the animal should lose the ability to navigate using spatial relationships. Richard Morris confirmed this in 1982 with his water maze, now one of the most widely used behavioral tests in neuroscience. Rats with hippocampal lesions could not learn to find a hidden platform using distal landmarks, while intact rats learned quickly. 4
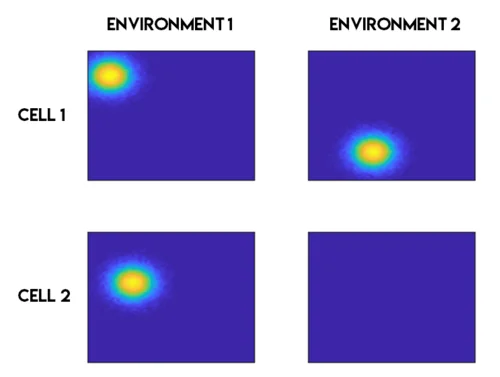
Place cells also proved more flexible than their initial description suggested. They undergo remapping — when an animal enters a familiar environment after some change (new color, new smell, new shape), many cells acquire completely different place fields or lose their fields entirely. Two environments that look similar to a human observer can evoke completely distinct patterns of place cell activity, as though the hippocampus is assigning each context its own unique map. This feature makes place cells well-suited for episodic memory: the spatial context of an event is stored alongside its content. 3
Grid cells: the coordinate system upstream
For three decades after 1971, a central question went unanswered: how do the place cells know where they are? The hippocampus receives its largest input from the entorhinal cortex, but entorhinal neurons showed only large, fuzzy place-like fields in early recordings. Then in 2005, Edvard Moser, May-Britt Moser, and their colleagues made a recording from the dorsomedial part of the entorhinal cortex — a part that previous studies had largely ignored — and found something astonishing.
Each recorded cell fired not at one location but at many, and those locations formed a precise hexagonal lattice covering the entire arena. Connecting the firing fields gave a tessellation of equilateral triangles: the vertex of every triangle was a firing field, and all triangles were the same size, the same orientation. This was a grid cell. 5

Grid cells fire regardless of landmarks, in darkness as well as light, and persist even after the environment has been disoriented. This suggested they were computing position by path integration — integrating the animal's own movement, speed, and direction over time. Cells in the same small patch of entorhinal cortex share the same grid spacing and orientation but differ in phase (which vertex aligns with which point in the room), and cells from more ventral parts of the entorhinal cortex have wider-spaced grids. The complete population of grid cells — with their varying spacings and phases — theoretically provides a unique firing-pattern label for every position in any reasonably sized environment.
The discovery of grid cells answered the question that place cells had posed: place cells know where they are because grid cells tell them. Grid cell inputs converge on hippocampal pyramidal neurons and, through mechanisms not yet fully resolved, collapse the repeating hexagonal signal into a single, location-specific place field. The navigational circuit finally had its architecture: grid cells provide the metric coordinate system; place cells store the allocentric map that the animal actually uses. 5
For this body of work — O'Keefe's place cells and the Mosers' grid cells — the 2014 Nobel Prize in Physiology or Medicine was awarded jointly to John O'Keefe, Edvard Moser, and May-Britt Moser.
What the cells reveal beyond space
The original cognitive map theory was explicitly spatial: the hippocampus is a spatial device. That framing has been challenged repeatedly, and the debate remains live.
Place cells in humans were recorded directly by Itzhak Fried and colleagues in 2003, using electrodes implanted in epilepsy patients. The findings held: individual hippocampal neurons fired selectively for specific virtual locations as patients navigated a simulated town. Human "place cells" responded to spatial position just as rodent ones did. 3
But place cells also respond to things that are not spatial. In 2017, Dmitriy Aronov and colleagues showed that hippocampal neurons in rats fire selectively for specific tones in a pitch-manipulation task — cells that would be "place cells" in a spatial context responded instead to position in a continuous sound-frequency space. The same architecture that maps physical location can apparently map other continuous variables too. This fits a broader "cognitive map" interpretation: the hippocampus represents any structured space — not just Euclidean geometry, but auditory space, social space, conceptual space. 3
The relationship between place cells and episodic memory points in the same direction. During sleep and rest, hippocampal place cells replay — they reactivate in rapid sequences that recapitulate the animal's recent path, or sometimes paths not yet taken (preplay). This replay, running at 10–20 times the normal speed, is thought to drive memory consolidation, transferring spatial and temporal information to the neocortex. The place cell is not just a position sensor; it participates in the process by which experience becomes memory.
Why this matters
Place cells gave neuroscience something rare: a clear, measurable neural correlate of a cognitive function that people actually care about. Before 1971, "where am I?" seemed like a question without a cellular answer. After 1971, you could point to specific neurons whose firing rate literally encodes the answer.
The downstream consequences are still unfolding. Alzheimer's disease — which devastates spatial navigation among its earliest symptoms — impairs place cell function in animal models before widespread neurodegeneration appears. Age-related cognitive decline shows up in degraded place cell precision. And because place cells connect to the RSC and PPA regions we have covered in recent articles, the navigation circuit described across this series now has its cellular foundation: the OPA and RSC process the visual and reference-frame information that flows into the hippocampus, where it is assembled into a map that can be read out by the animals — and by us — as "I am here."
The 1971 paper was 13 lines of results. The 2014 Nobel Prize was the delayed receipt for what those 13 lines had opened.
Landmark paper: O'Keefe, J. & Dostrovsky, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Research, 34(1), 171–175. PMID: 5124915
Course connection: MIT 9.13 covers the hippocampus and spatial navigation as part of its module on navigation and spatial cognition. Place cells and grid cells are central examples in Nancy Kanwisher's lectures on how the brain represents space and supports memory.
이 콘텐츠를 둘러싼 관점이나 맥락을 계속 보강해 보세요.